ARTÍCULO ORIGINAL
Revista Científico estudiantil Ciencias Forestales y Ambientales.
Enero-junio 2018; 3(1):89-113
![]()
Habitats modelling in Pinus caribaea var. caribaea Barrett and Golfari and Pinus tropicalis Morelet
Yordani Crespo Martínez
Universidad de Pinar del Río "Hermanos Saíz Montes de Oca". Cuba.
Recibido: 22 de enero 2018.
Aprobado: 26 de febrero 2018.
RESUMEN
Con el objetivo de modelar los hábitats potenciales de Pinus caribaea var. caribaea y Pinus tropicalis, se realizó un análisis bajo el algoritmo de Máxima Entropía, con datos de presencia de las especies y datos de variables bioclimáticas de un modelo climático global y con una base de datos observados (1981-2010). Se georreferenciaron 258 y 1 381 puntos como datos de presencia para P. caribaea y P. tropicalis. El régimen de precipitaciones es el más importante para ambas especies. Las áreas idóneas para el desarrollo de las especies se corresponden con las áreas más densamente ocupadas por los pinares naturales, no así con las plantaciones. El área potencial aproximada, donde la probabilidad de presencia sea superior a 0.6 en P. caribaea, fue de 26 900 ha. para el modelo de la serie observada, mientras que para P. tropicalis fue 88 200 ha. Los hábitats potenciales se concentran en la zona occidental de Cuba, fundamentalmente en la provincia de Pinar del Río.
Palabras clave: P. tropicalis; P. caribaea; modelación de hábitats potenciales; MaxEnt.
ABSTRACT
The aims of this study was to model the potencial habitats of P. caribaea and P. tropicalis using the Max Ent algorithm based on the relation between presence data and climatological data from a global climatic model or observed climatic serie (1981-2010). 258 and 1 381 natural stand were georeferenced as a presence data. The precipitation regimen was most important factor for both species. The potential suitable areas agree with the current densest areas for both species, but not for the planted areas in P. caribaea. The approximated potential areas with a higher presence probability (0.6) in P. caribaea was 26 900 ha for the observed data; whereas for P. tropicalis was around 88 200 ha. Besides, these models clearly shown that the potential habitats laid on the western of Cuba, exclusively in Pinar del Río
Key words: P. tropicalis; P. caribaea; modelling potential habitats; MaxEnt.
INTRODUCCIÓN
Ya sea por el cambio climático, que experimenta hoy en día nuestro planeta, o por el desarrollo de la agricultura, la que, desde el mismo momento de su surgimiento, ha modificado en gran escala la cobertura vegetal natural de todos los continentes sobre la tierra, los hábitats de muchas especies vegetales y animales han cambiado en mayor o menor medida por los cambios en las oscilaciones climáticas, Saunders et al. (1991); Fong, (2008).
El género Pinus incluye 111 especies, Richardson et al., (1998); Geada et al., (2002), Gernandt et al., (2005); Piloto, (2015); más de 30 viven al sur del Trópico de Cáncer y durante su evolución han experimentado grandes cambios que les ha permitido adaptarse. Esto ha favorecido que este género sea un buen modelo para estudiar cómo afectarían los cambios, en el clima su distribución. Las especies más interesantes son las que viven en el área del Caribe, donde existen barreras físicas que limitan su distribución Whittaker y Fernández, (1998), Jardón et al., (2011).
En la isla de Cuba, habitan cuatro especies endémicas de pino: Pinus tropicalis Morelet, P. caribaea Morelet var. caribaea Barret y Golfari, Pinus cubensis Griseb y Pinus maestrensis Bisse; de ellas, dos, en la zona occidental y las dos últimas, en la oriental, aunque solamente P. tropicalis tiene un origen euroasiático.
Estas especies son de suma importancia, económica y ecológica, pues ocupan grandes extensiones de tierra en el archipiélago y, por tanto, en los bosques de pinos cubanos, se puede encontrar una gran cantidad de recursos madereros y no madereros y una gran variedad de especies animales y vegetales las cuales se desarrollan en este tipo de formación boscosa. Las provincias de Pinar del Río e Isla de la Juventud cuentan con grandes extensiones de pinares.
Tanto P. tropicalis como P. caribaea son las principales especies utilizadas en la producción de madera aserrada, siendo, además, entre las cuatro especies priorizadas en los planes de reforestación hasta el 2030. Para garantizar el cumplimiento de esos planes, se hace necesario conocer las áreas que posean las condiciones más idóneas para su desarrollo. Generalmente, dicha información no se conoce con exactitud, por lo que contar con un Sistema de Información Geográfica (SIG), que facilite este proceso, sería de gran utilidad.
El modelado ecológico se ha convertido en una herramienta muy potente que permite generar modelos de distribución de especies (MDE) los que son una alternativa que brinda la información necesaria sobre las condiciones climáticas y áreas, en las cuales, determinada especie se puede desarrollar. Existen dos alternativas para estimar los patrones de distribución de hábitats potenciales, ya sea intuitivamente, o bien al relacionar localidades de los registros con variables, mediante el uso de modelos empíricos predictivos de distribución potencial, Ávila et al., (2014). Estos modelos empíricos o MDE tienen por objeto predecir áreas que describan las condiciones ambientales adecuadas para la supervivencia de las especies; es decir, la distribución potencial o nicho fundamental, Guisan y Thuiller, (2005); Ledig et al., (2010); Ledig, (2012).
En general, estos métodos de modelación combinan datos de localidades georreferenciadas de las especies, donde ha sido confirmada su presencia con variables medioambientales para crear un modelo de requerimientos de la especie de acuerdo con las variables examinadas, Anderson et al., (2003).
Existen varios algoritmos matemáticos para el modelado de hábitats potenciales como pueden ser: BIOCLIM (Envolventes Bioclimáticas), MLJ (Modelado Lineal Generalizado, Domian (Domian Modeling), GARP (Genetic Algorithm for Rule.set Prediction) y MAG (Modelado Aditivo Generalizado) o los métodos de aprendizaje cibernético como MaxEnt (Algoritmo de Máxima Entropía).
El uso de estas aplicaciones combinadas con una herramienta SIG puede generar mapas de distribución potencial de especies, lo que resulta fundamental para evaluar las condiciones ambientales de una localidad o sitio para el desarrollo de una determinada especie. Narkis, (2012).
En Cuba, no se han realizado muchos estudios sobre la base de MDE y solo se cuenta con pocos estudios en animales (anfibios, gruyas) y, hasta el momento, se documenta un estudio sobre modelación en P. tropicalis y P. caribaea. (Miranda et al., (2016).
Esta investigación partió de la necesidad de conocer la proyección de nuevas áreas potenciales para ambas especies y así ofrecer indicaciones precisas sobre disponibilidad de áreas. Por lo que tuvo como objetivo: modelar los hábitats potenciales de P. caribaea y P. tropicalis.
MATERIALES Y MÉTODOS
Ubicación del área de estudio
El estudio se realizó para parte de la región occidental de Cuba (Figura 1) que está entre las coordenadas 22°242 443 N y 83°402 193 O. Los datos de presencia se tomaron a partir de datos de distribución de los pinares naturales, tomados de las bases de datos SIFOMAP 1.5.2 (Sistema Informativo Automatizado para la Gestión Forestal) correspondientes a las empresas de la región.
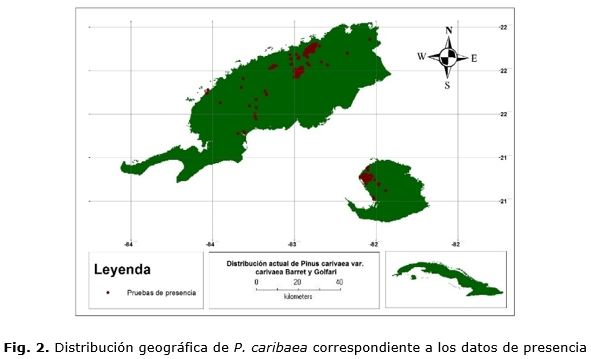
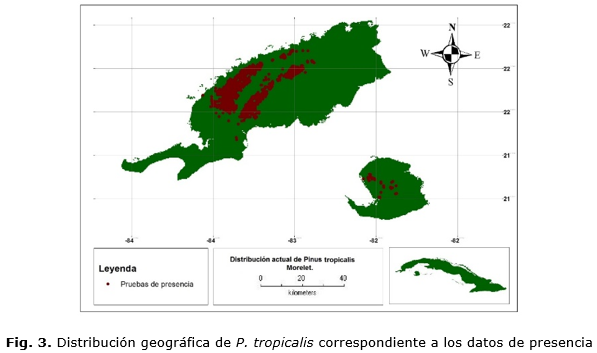
Para la confección de la base de datos, se georreferenciaron 258 y 1381 puntos de presencia para P. caribaea (Figura 2)y P. tropicalis (Figura 3) respectivamente, bajo el supuesto que los pinares naturales de ambas especies representan las condiciones climáticas necesarias para su desarrollo.
Variables bioclimáticas
Las variables bioclimáticas (Tabla 1) fueron tomadas de dos formas: una, utilizando la base de datos del portal de Earth System Grid (ESG) de WorldClim (www.worldclim.org) para el clima observado (1960-1990) bajo el Modelo Climático Global CCM3 y la segunda de la serie histórica de 30 años de las estaciones meteorológicas de Pinar del Río e Isla de la Juventud. Con la información de estos datos climáticos, se obtienen 17 y 18 capas RASTER en formato ASCII.

Confección del modelo
Para la elaboración del modelo, se empleó el algoritmo MaxEnt (Máxima Entropía) ver. 3.3.3 Phillips et al., (2004); Phillips y Dudik, (2010), Elith et al., (2011), utilizando la información de las variables bioclimáticas y los datos de presencia.
MaxEnt modela la distribución geográfica de las especies, determinando la relación entre las variables bioclimáticas y los registros de las especies Guisan y Thuiller, (2005); Palma y Delgadillo, (2007). El resultado de la modelación con MaxEnt revela la probabilidad relativa de distribución de una especie en todas las cuadrículas o celdas en el espacio geográfico definido, en el cual un valor de alta probabilidad, asociado a una cuadrícula particular, indica la probabilidad de la misma, de tener condiciones ambientales favorables para la especie modelada, Elith et al., (2006).
La configuración del modelo se realizó con la función logística, por ser la más sencilla de conceptualizar, ya que proporciona un estimado entre cero y uno de probabilidad de presencia, Phillips et al., (2006); para ellos, se empleó el 75 % de los datos para entrenamiento y el 25 % para validación. En la calibración del modelo, se especifican 5000 interacciones y el límite de convergencia se fijó en 0.00001 (valor por defecto), lo que garantiza la convergencia del algoritmo.
La capacidad predictiva del modelo se determinó con la técnica de evaluación del área bajo la curva (AUC), de la curva operacional (ROC). El AUC representa la relación entre el porcentaje de presencia correctamente predicha (sensibilidad) y, uno menos el porcentaje de ausencias correctamente predichas (especificidad). El AUC mide la capacidad del modelo para clasificar correctamente a una especie como presente o ausente, (Hanley y McNeil, (1982); Palma y Delgadillo, (2007). Los valores de AUC pueden ser interpretados como la probabilidad de que, cuando se seleccione al azar un punto de presencia y uno de ausencia, el punto de presencia tendrá un valor mayor de idoneidad o probabilidad sobre el modelo.
De acuerdo con Araújo y Guisan, (2006); Palma y Delgadillo, (2007) una guía general para la clasificación de la precisión del modelo, tomando en cuenta el AUC, es:
0.50-0.60 = insuficiente;
0.60-0.70 = pobre;
0.70-0.80 = promedio;
0.80-0.90 = bueno;
0.90 a 1 = excelente.
Para evaluar las variables bioclimáticas que más contribuyeron al modelo, se realizó la prueba de Jackknife. Esta información es de suma importancia para evidenciar los requerimientos ecológicos de cada especie.
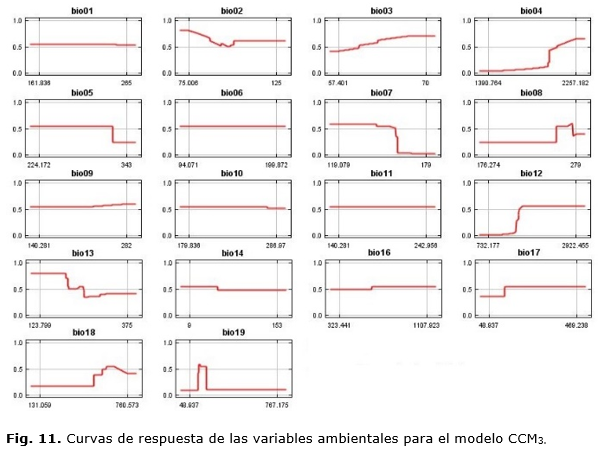
Adicionalmente, se realizaron las curvas de respuesta para describir cómo cada variable afecta de forma individual la predicción.
Los mapas resultantes fueron analizados y procesados con el Sistema de Información Geográfica para Análisis de la Distribución de Especies DIVA-GIS ver 7.3.0 (Hijmans et al., (2004).
RESULTADOS Y DISCUCIÓN
Modelación de hábitat potencial
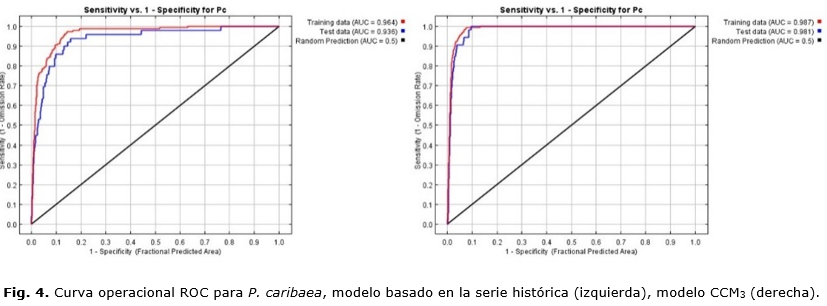
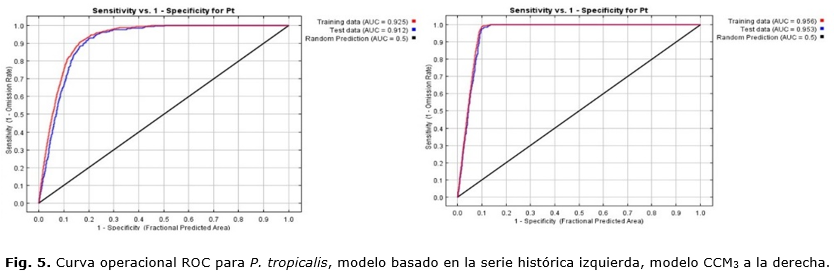
Los modelos generados con el MaxEnt para P. caribaea y P. tropicalis fueron capaces de proyectar el área potencial de la especie. El AUC de la curva fue mayor que 0.9, tanto para los datos de entrenamiento como los de validación (datos de prueba); esto significa que el modelo puede predecir con alto nivel de especificidad y sensibilidad el área potencial. De acuerdo con Araújo y Guisan, (2006); Ávila et al., (2014) y Palma, (2007), valores por encima de 0.9 son considerados como muy bueno en el desempeño del modelo.
En modelo generado con el CCM3, en relación con el de la serie observada, para P. caribaea y P. tropiaclis obtiene un mayor valor de AUC (Figura 4 y 5); sin embargo, esto no significa que el modelo sea de mayor resolución y capaz de detectar las variaciones a menor escala espacial. De ahí que hay que ser muy cuidadoso cuando se interpretan los resultados, en especial para las islas, donde los modelos globales no permiten la detección de las variaciones espaciales pues dependen del número de estaciones que se utilicen como referencia, (Karmalkar et al., (2013); Silva et al., (2015). Por tanto, siempre que sea posible debe modelarse con datos observados de series históricas de más de 30 años para ser más fiable en la elaboración de la línea base en estudios de impacto del cambio climático (Ledig et al., (2010).
Altos valores de AUC pueden estar ligados a la selección de un entorno ambiental disponible mayor que el de las áreas de prevalencia de la especie Cobos, (2016), al incremento del área de calibración Jiménez, (2008) y diferencias mayores entre registros de presencia distantes en relación con los más cercanos VanDerWal et al., (2009) como cuando se trabaja con modelos globales. No obstante, Miranda et al., (2016), con otra proporción de datos prueba y entrenamiento menor, obtuvieron un resultado similar a este (0.917).
Lo que parece una generalidad es que para especies arbóreas los valores del desempeño de los modelos se encuentran por encima de 0.8 (Araujo y Guisan, (2006); Kumar y Stohlgren, (2009); Contreras et al., (2010); Ávila et al., (2014). Esto pudiera estar dado a que la estrecha correlación, que se establece entre las variables bioclimáticas (variables meteorológicas) y la presencia de la especie, es más dependiente de ellas que de otras variables ambientales, como es en el caso de los animales.
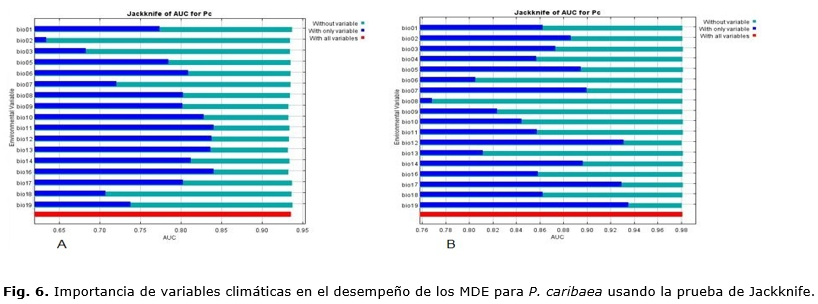
Contribución de las variables bioclimáticas a los modelos para P. caribaea
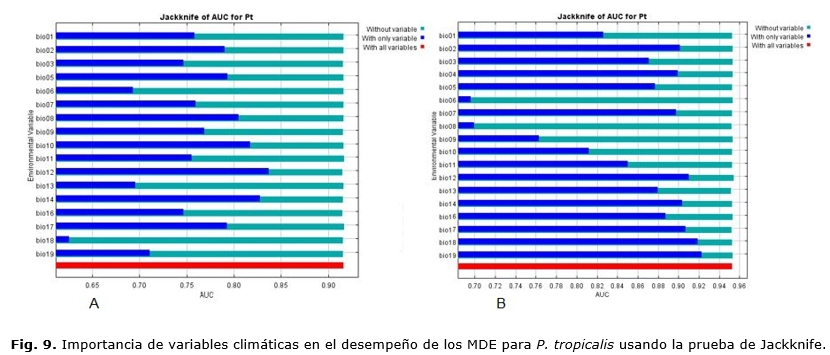
La prueba de Jackknife facilita la identificación de las variables más importantes al modelo, el MaxEnt, de manera gráfica; muestra cuál variable contribuye más al modelo.
En este caso, hubo diferencias en las variables más importantes a cada modelo. El modelo basado en serie observada (Figura 6 A) mostró que las variables relacionadas con régimen de precipitación bio16 (precipitación del trimestre más lluvioso), bio12 (precipitación media anual), bio13 (precipitación del período más lluvioso) y en segundo lugar la bio11 (temperatura media del trimestre más frío) fueron más importantes para la especie. Sin embargo, para el modelo generado con el CCM3 (Figura 6 B), solamente aquellas referidas a las precipitaciones como: bio 12 (precipitación media anual), bio17 (precipitación del trimestre más seco) y bio19 (precipitación del trimestre más frío) fueron importantes.
De manera general, los modelos son congruentes al señalar que el régimen de precipitaciones es el que más determina la presencia de la especie. Esto es de gran importancia, si se conoce que Cuba ha documentado las variaciones en su clima, expresadas en la reducción de las precipitaciones y el aumento de la temperatura media, Centella et al., (2001); Álvarez et al., (2014); Cutié et al., (2013), por lo que, entonces, podría inferirse que la especie será seriamente afectada en el futuro, de continuar esta tendencia.
Curvas de respuesta de las variables bioclimáticas para P. caribaea
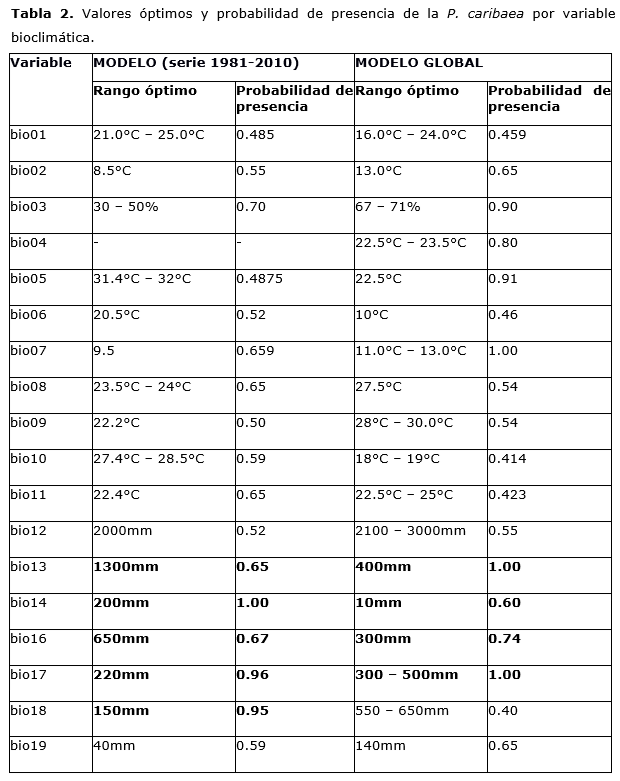
La tabla 2 muestra los rangos en que cada variable climática alcanza valores mayores de probabilidad de presencia para estos valores y que, por tanto, permite establecer los rangos climáticos de la especie y el umbral al cual se vería afectada su presencia (Figura 7 y 8) de manera independiente para cada variable.
Las figuras 7 y 8 muestran gráficamente cómo las variables ambientales, de forma independiente, afectan las predicciones de MaxEnt, o sea, la forma en que la predicción logística cambia en la medida que cada variable ambiental es variada, manteniendo todas las otras variables ambientales en su valor promedio. El eje Y representa la probabilidad de presencia y el eje X representa los valores de cada variable.
Se puede apreciar cómo, para el modelo basado en la serie histórica, no se afecta la predicción en el caso de bio01, bio02, bio05, bio10, bio11 y bio18 pues la misma se mantiene constante en la medida que el valor de cada una de estas variables es cambiado pues ellas no van a ejercer gran influencia en la predicción. Las variables que menos afectan la predicción, en el modelo CCM3, son: bio01, bio02, bio05, bio10 y bio11. Para ambos modelos, se define que las variables que menos afectan la predicción son aquellas relacionadas con las temperaturas. Esto puede deberse a que el área de estudio comprende una isla relativamente estrecha por lo que no existen grandes diferencias en cuanto al valor de las temperaturas.
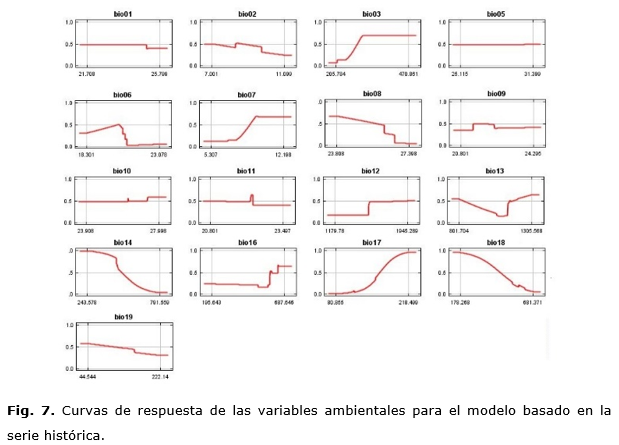
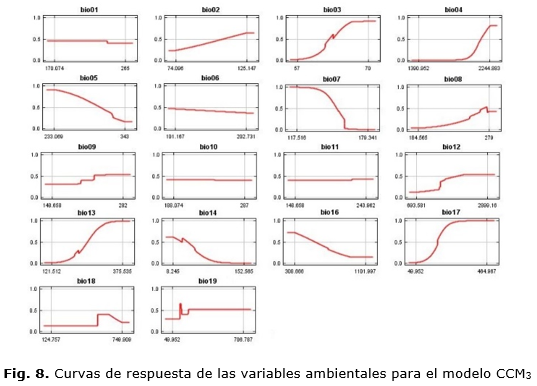
Ambos casos son congruentes al mostrar que la especie podría comprometer su presencia al disminuir el régimen de precipitaciones anuales y, si aumenta este en el periodo más seco, afectaría su presencia. Planos et al., (2013), reportan que, durante los últimos 30 años, se ha observado una ligera inversión en el patrón de lluvias lo cual tendría, entonces, un efecto negativo en la especie de continuar o intensificarse.
Contribución de las variables bioclimáticas a los modelos para P. tropicalis
De acuerdo con los resultados de la prueba de Jackknife (Figura 9 A) se determinó que las variables que más influyeron para P. tropicalis, en el caso de modelo basado en la serie histórica, fueron: bio12 (precipitación anual), bio14 (precipitación del periodo más seco) y bio10 (temperatura media del trimestre más cálido). En este caso, la especie responde a variaciones no solo de las precipitaciones, sino de la temperatura.
Para el modelo CCM3 (Figura 9 B), solamente las variables relacionadas con el régimen de precipitaciones fueron importantes: bio19 (precipitación del trimestre más frío), bio18 (precipitación del trimestre más cálido), bio12 (precipitación anual), bio17 (precipitación del trimestre más seco). Esto pudiera deberse a que el modelo no expresa variación en las variables de temperatura en las áreas de presencia.
En ambas especies y ambos modelos, se definió que las precipitaciones juegan un papel fundamental en la distribución de las especies Se identificó que P. tropicalis soporta regímenes de precipitaciones más secos y este puede desarrollarse con facilidad en condiciones donde haya déficit de agua. Para el caso de P. caribaea, las precipitaciones deben ser más abundantes que para P. tropicalis, aunque esto no, precisamente, determina que esta no sobreviva en condiciones de poca agua, si sugiere que el estrés hídrico puede llevar a la especie a un desarrollo lento y poco eficiente.
Téllez et al., (2005) y Ávila et al., (2014) argumentan el uso del modelado bioclimático como herramienta para el manejo forestal de cuatro especies de Pinus para México pues facilita la determinación de los rangos climáticos de cada especie y que, además, pueden usarse otras variables ambientales, si se dispone de información como la pendiente, el suelo, entre otras.
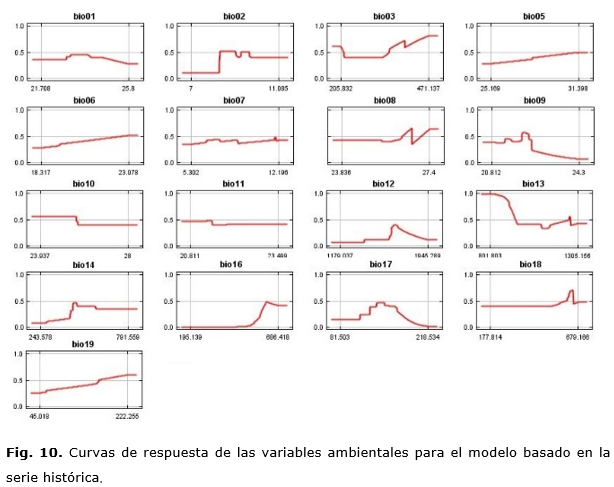
Curvas de respuesta de las variables bioclimáticas para P. tropicalis
La Tabla 3 muestra los rangos óptimos de cada variable climática para P. tropicalis, así como la probabilidad de presencia para estos valores. Para esta especie, las variables que más influyen corresponden también al régimen de precipitaciones; pero en este caso posee una mayor flexibilidad en cuanto a los valores óptimos de estas.
Tabla 3. Valores óptimos y probabilidad de presencia de P. tropicalis por variable climática.
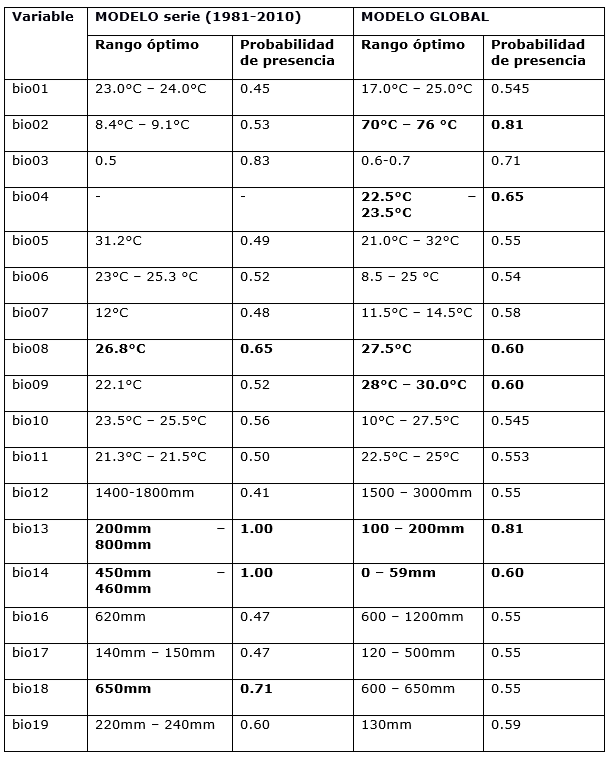
Las curvas de respuesta muestran que, si las precipitaciones medias anuales están entre 1400 a 1800 mm., la respuesta a la presencia es positiva hasta 1900 mm. Si la temperatura media del trimestre más cálido aumenta ligeramente, no afectará grandemente la presencia de la especie.
Distribución de hábitats potenciales
Los modelos de distribución de hábitats potenciales de P. caribaea y P. tropicalis resultaron de la combinación de las variables para cada uno de los modelos. Los mapas proporcionan un estimado de cero a uno de probabilidad a la presencia de la especie Phillips et al., (2004); Phillips y Dudik, (2010); Elith et al., (2011).
Distribución potencial para la especie P. caribaea basado en serie histórica y el modelo climático global
Los mapas resultantes de la distribución potencial para la especie P. caribaea indican que la mayor probabilidad de presencia se encuentra en áreas muy similares a las que ocupa la especie en la actualidad, aunque solo para los pinares naturales. Sin embargo, las plantaciones de esta se encuentran en lugares donde según el modelo no hay condiciones climáticas para el desarrollo de la especie.
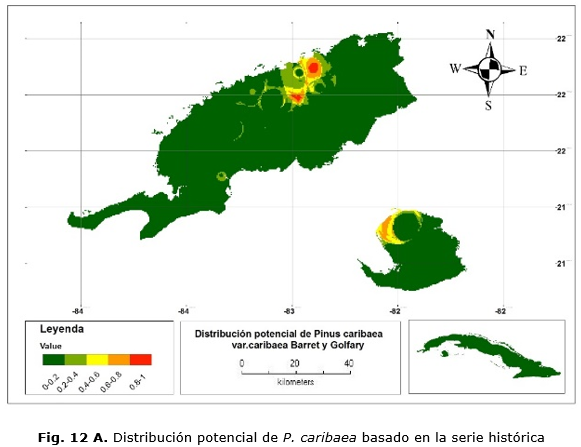
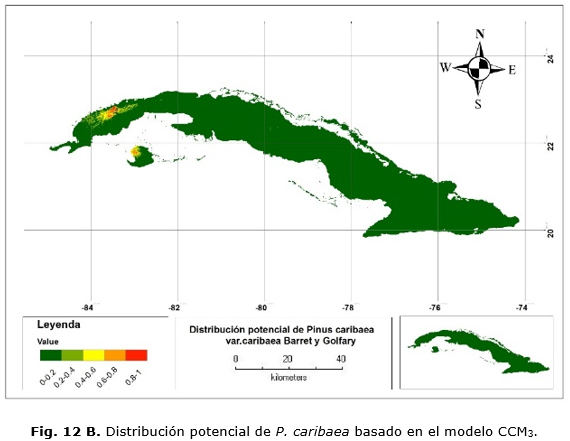
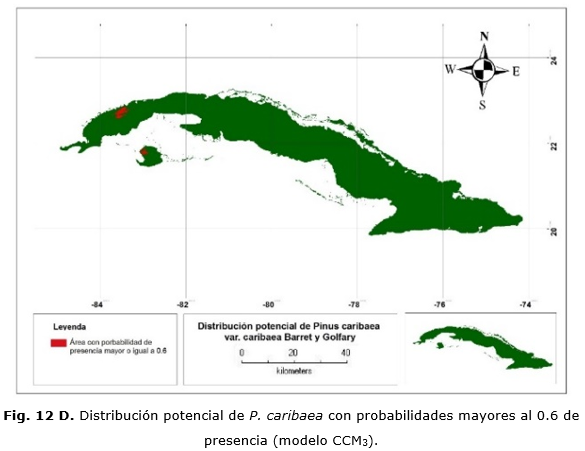
El modelo basado en la serie histórica muestra que las áreas de distribución de la especie se van a encontrar principalmente en las áreas que comprenden a las localidades de La Palma, San Andrés y la región noroeste de la Isla de la Juventud, no así en los territorios de Mantua, Guane, San Juan y Martínez, Minas de Matahambre (Figura 12 A). En el caso del modelo global, el área predicha abarca aproximadamente el mismo territorio, solo que el área se extiende un poco más al oeste (Figura 12 B).
Aunque ambos modelos sitúan las áreas de distribución en localidades similares a las actuales, estos discrepan en cuanto a las probabilidades de presencia de la especie en estas áreas. En el modelo regional, la probabilidad de presencia en la gran mayoría del territorio es de valores que oscilan entre 0.4 y 0.8 mientras que el modelo global arrojó valores para la mayor parte del territorio, comprendidos entre 0.6 y 1.
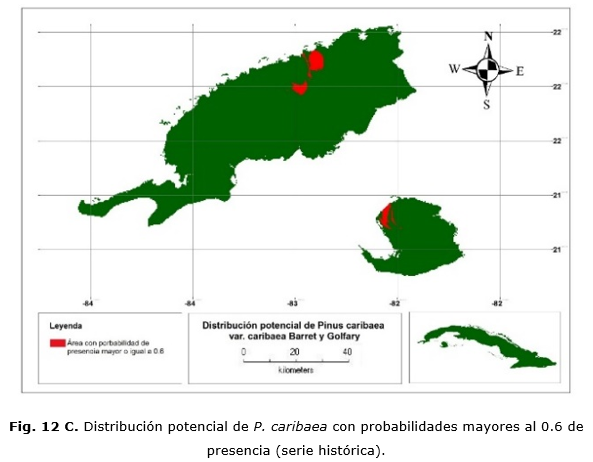
Las Figuras 12 C y 12 D muestran las áreas donde se concentran las mayores probabilidades de presencia y que representarían las condiciones óptimas para su desarrollo. En cualquier caso, esto abarca 26 900 ha.; de ellas, 15 700 ha. en la provincia de Pinar del Río y 9 200 ha. en Isla de la Juventud. Con el modelo CCM3, existe una sobrestimación de 62 200 ha.; 55 800 y 6 400 ha. para Pinar del Río e Isla de la Juventud, respectivamente.
Distribución potencial para la especie P. tropicalis basado en serie histórica y el modelo climático global
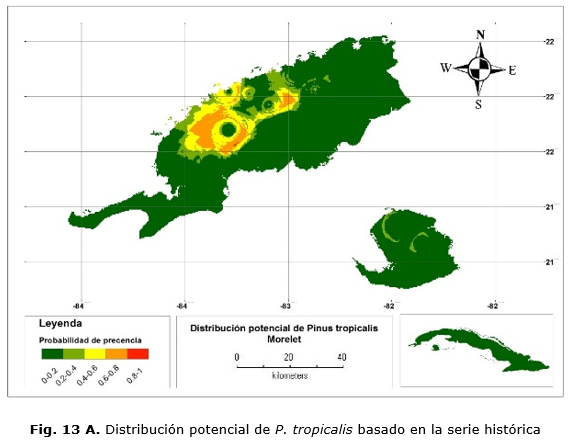
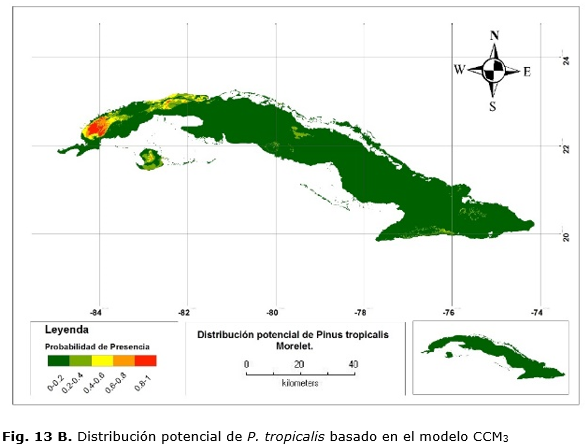
Los mapas muestran que las áreas más idóneas para la distribución de P.t, al igual que el anterior, se concentran en áreas donde la especie se encuentra actualmente formando pinares monotípicos. En este caso, se encuentran en estas áreas, además, plantaciones P. caribaea, donde no son favorables estas condiciones para la especie.
Por su parte, el modelo generado con la serie observada, históricamente muestra que las áreas de distribución abarcan los territorios que comprenden las localidades de los municipios Mantua, Guane, San Juan y Martínez, Minas de Matahambre y Viñales y no así en Isla de la Juventud (Figura 13 A).
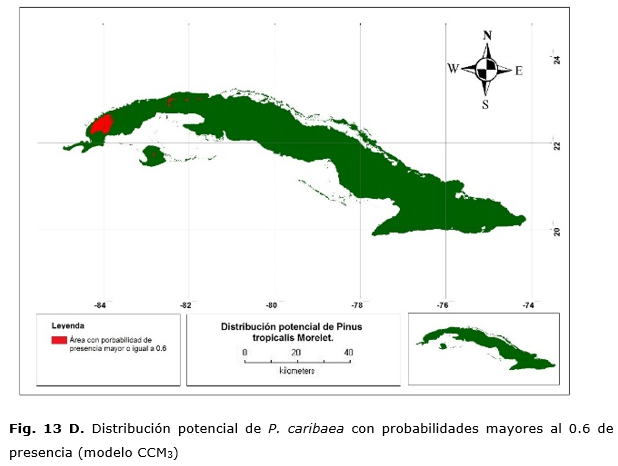
Las áreas predichas por el modelo CCM3 indican que el área de mayor probabilidad de presencia de la especie se encuentra en estos municipios, pero, además, muestra una pequeña área perteneciente al norte de la provincia de Artemisa y el centro de Isla de la Juventud (Figura 13 B).
Existe congruencia entre los modelos para las áreas predichas, aun siendo uno espacialmente más grande y con menos resolución. Por lo tanto, las condiciones más favorables se encuentran para las dos especies en Pinar del Río.
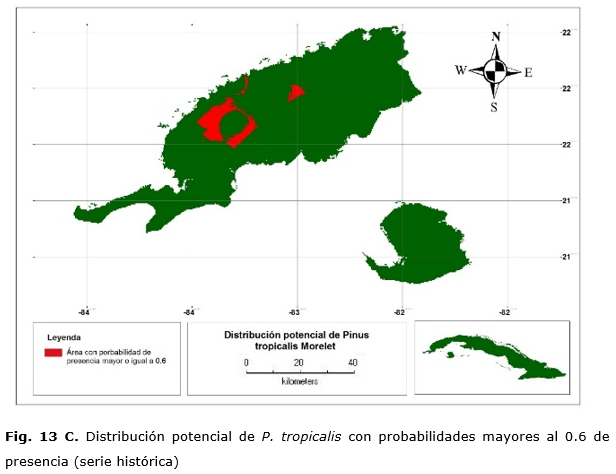
Las Figuras 13 C y 13 D muestran las áreas donde se concentran las mayores probabilidades de presencia (e» 0.6) y que representarían las condiciones óptimas para su desarrollo. En cualquier caso, esto abarca 88 200 ha. en Pinar del Río ya que para la Isla de la Juventud no hay probabilidades que superen 0.6. Sin embargo, con el modelo CCM3, predice de 162 850 ha., 145 550, 2 600 y 14 700 ha. para Pinar del Río, Isla de la Juventud y Artemisa, respectivamente. Muchas de estas áreas se encuentran fuera de la distribución actual de la especie por la existencia de barreras edáficas para su desarrollo como lo son los suelos cársicos o derivados de serpentinas y otros son suelos agrícolas, que antes estuvieron con una formación de bosque semideciduo Borhidi, (1996). Por lo que el modelo generado con los datos del CCM3 tiene imprecisiones para estimar los hábitats.
Los modelos de distribución de hábitats potenciales realizados para ambas especies mostraron que tienen áreas potenciales en diferentes direcciones, más al noroeste para P. caribaea y al oeste P. tropicalis, donde esta última es la que posee mayores áreas potenciales para su desarrollo.
El modelo realizado con los datos de la serie observada es capaz de predecir las áreas de manera más eficaz, debido a una mejor resolución, además que se originó sobre datos climáticos de mayor cantidad de estaciones, lo que le brinda al modelo la capacidad de describir las condiciones climáticas de las áreas de desarrollo de la especie, de manera más exacta. Por otra parte, con el modelo global se pudieron detectar áreas para el desarrollo de la especie que no necesariamente se encuentran ubicadas en la provincia de Pinar del Río.
CONCLUSIONES
Las variables bioclimáticas que más influyen en la distribución de pinares de P. caribaea son el régimen anual de precipitaciones y, para P. tropicalis, son la precipitación anual y la temperatura en el período más cálido.
P. caribaea requiere valores de precipitaciones anuales mayores a 1 800 mm., en el período más seco de 200 mm.; mientras que P. tropicalis precisa de 1 600 hasta 1 800 mm. y precipitaciones de 400-450 mm., en el período más seco.
El modelo prevé que P. caribaea tiene mayor cantidad de hábitats potenciales hacia el noreste de Pinar del Río y noroeste de la Isla de la Juventud y P. tropicalis se concentra principalmente hacia el oeste de Pinar del Río; en el resto del territorio no se detectaron hábitats potenciales para las especies.
REFERENCIAS BIBLIOGRÁFICAS
Álvarez-Brito, A., Mercadet-Portillo, A. (2014). El sector forestal cubano y el cambio climático. Instituto de Investigaciones Agroforestales. Ministerio de la Agricultura. Anales de la Academia de Ciencias de Cuba, 4(2), 11.
Anderson, R. P., Lew, D., & Townsend Peterson, A. (2003). Evaluating predictive models of species distributions: criteria for selecting optimal models. Ecological Modelling, 162(3), 211-232. https://doi.org/10.1016/S0304-3800(02)00349-6.
Araújo, M. B., & Guisan, A. (2006). Five (or so) challenges for species distribution modelling. Journal of Biogeography, 33(10):16771688. https://doi.org/10.1111/j.1365-2699.2006.01584.x.
Ávila-Coria, R., Villavicencio García, R., Ruiz Corral, J. A. (2014). Distribución potencial de Pinus herrerae Martínez en el Occidente del estado de Jalisco. Revista Mexicana de Ciencias Forestales, 5(24), 93-108.
Barbolla, L. J., Delgado-Valerio, P., Geada-López, G., Vázquez-Lobo, A., Piñero, D. (2011). Phylogeography of Subsection Australes pines in the Caribbean basin. Annals of Botany, 107(2), 229-241. https://doi.org/10.1093/aob/mcq232
Borhidi, A. (1996). Phytogeography and vegetation ecology of Cuba. 28(1). Budapest: Akademiai kiado. Recuperado a partir de https://link.springer.com/content/pdf/10.1007%2FBF02853198.pdf
Centella-Artola, A., Llanes-Regueiro, J., Paz-Castro, L., López, C., Limia-Martínez, M. (2001). Primera Comunicación Nacional a la Convención Marco de las Naciones Unidas sobre Cambio Climático, República de Cuba (p. 166). La Habana, Cuba: Instituto de Meteorología-CUBAENERGIA. Recuperado a partir de http://unfccc.int/resource/docs/natc/cubnc1.pdf
Cobos Cobos, M. E. (2016, marzo 11). Posibles implicaciones del cambio climático sobre la distribución de las especies del género Peltophryne (Anura: Bufonidae) en Cuba (Tesis de maestría). La Habana, Cuba. Recuperado a partir de http://repositorio.educacionsuperior.gob.ec/handle/28000/2470, http://repositorio.educacionsuperior.gob.ec/bitstream/28000/2470/1/T-SENESCYT-01321.pdf
Contreras-Medina, R., Luna Vega, I., Ríos Muñoz, C. A. (2010). Distribución de Taxus globosa (Taxaceae) en México: Modelos ecológicos de nicho, efectos del cambio del uso de suelo y conservación. Revista Chilena de Historia Natural, 83, 421-433.
Cutié-Cancino, V., Lapinel-Pedroso, B., Gonzáles-Rodríguez, N., Perdigón-Morales, J., Fonseca-Rivera, C., Gonzáles-García, I. (2013). La sequía en Cuba, un texto de referencia. Monografía. Proyecto 1/OP-15/GEF. La Habana, Cuba: Instituto de Meteorología.
Elith, J., Graham, C. H., Anderson, R. P., Dudík, M., Ferrier, S., Guisan, A., Huettmann, F. (2006). Novel methods improve prediction of species' distributions from occurrence data, 29(2), 129-151. https://doi.org/10.1111/j.2006.0906-7590.04596.x
Elith, J., Phillips, S. J., Hastie, T., Dudík, M., Chee, Y. E., Yates, C. J. (2011). A statistical explanation of MaxEnt for ecologists. Diversity and Distributions, 17(1), 4357. https://doi.org/10.1111/j.1472-4642.2010.00725.x
Fong Grillo, A. (2008). Distribución y conservación de los anfibios de los macizos montañosos de la región oriental de Cuba (Tesis doctoral). Universidad Complutense de Madrid Facultad de Ciencias Biológicas, biología vegetal I, Madrid, España.
Geada López, G., Kamiya, K., Harada, K. (2002). Phylogenetic relationships of Diploxylon pines (Subgenus Pinus) based on plastid sequence data. International Journal of Plants Science, 163(5), 737-747. https://doi.org/10.1086/342213
Gernandt, D. S., Geada López, G., Ortiz García, S., Liston, A. (2005). Phylogeny and classification of Pinus. International Association for Plant Taxonomy (IAPT), Taxon, 54(1), 29-42. https://doi.org/10.2307/25065300
Guisan, A., Thuiller, W. (2005). Predicting species distribution: offering more than simple habitat models. Ecology Letters, 8(9), 9931009. https://doi.org/10.1111/j.1461-0248.2005.00792.x
Hanley, J. A., McNeil, B. J. (1982). The meaning and use of the area under a Receiver Operating Characteristic (ROC) curve. Radiology, 143(1), 29-36.
J. Hijmans, R., Guarino, L., Bussink, C., Mathur, P., Cruz, M., Barrantes, I., Rojas, E. (2004). Sistema de Información Geográfica para el Análisis de Datos de Distribución de Especies. Universidad de California. USA: DIVA-GIS, Versión 4. Recuperado a partir de http://www.diva-gis.org/docs/DIVA-GIS4_manual_Esp.pdf
Jiménez-Valverde, A., M. Lobo, J., Hortal, J. (2008). Not as good as they seem: the importance of concepts in species distribution modelling. Diversity and Distributions, 14(6), 885890. https://doi.org/10.1111/j.1472-4642.2008.00496.x
Karmalkar, A. V., Taylor, M. A., Campbell, J., Stephenson, T., New, M., Centella, A., Charlery, J. (2013). A review of observed and projected changes in climate for the islands in the Caribbean. (edited by H. Diaz). Atmósfera, 26(2). Recuperado a partir de http://www.revistascca.unam.mx/atm/index.php/atm/article/view/36417
Kumar, S., Stohlgren, T. J. (2009). Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. Journal of Ecology and Natural Environment, 14, 94-98.
Ledig, F. T. (2012). Climate change and conservation. Acta Silvatica et Lignaria Hungarica, 8, 5774.
Ledig, F. T., Rehfeldt, G. E. C., Sáenz-Romero, Flores-López, C. (2010). Projections of suitable habitat for rare species under global warming scenarios. American Journal of Botany, 97(6), 970-987.
Miranda-Sierra, C. A., Geada-López, G., Sotolongo-Sospedra, R. (2016). Modelación de la distribución potencial de Pinus tropicalis en el Occidente de Cuba. Avances, 18, enero-marzo (1), 28-35.
Narkis-Morales, S. (2012). Modelos de distribución de especies: Software Maxent y sus aplicaciones en Conservación. Revista Conservación Ambienta, 2(1), 1-5.
Palma-Ordas, S., Delgadillo-Rodríguez, J. (2007). Distribución potencial de ocho especies exóticas de carácter invasor en el estado de Baja California, México. Botanical Sciences, 92(4), 587-597.
Phillips, S., Dudik, M. (2010). Una breve guía didáctica sobre MaxEnt. AT&T. Princeton University y el Centro para la Biodiversidad y Conservación del Museo Americano de Historia Natural. Recuperado a partir de http://manualzz.com/doc/21178397/una-breve-gu%C3%ADa -did%C3%A1ctica-sobre-maxent—por-steven-phillip...
Phillips, S. J., Dudik, M., Schapire, R. E. (2004). A maximum entropy approach to species distribution modeling. En Proceedings of the 21st International Conference on Machine Learning (pp. 655-662). Banff, Canadá. Recuperado a partir de https://dl.acm.org/citation.cfm?id=2856767&picked=prox
Phillips, S. J., Dudik, M., Schapire, R. E. (2006). Maximum entropy modelling of species geographic distributions. Ecological Modelling, 190(3-4), 231-259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Piloto, J. A. (2015). Variación anatómica de Pinus caribaea var. caribaea Barrett y Golfari y Pinus tropicalis Morelet (Tesis de Diploma). Pinar del Río.
Planos Gutiérrez, E. O., Guevara Velazco, A. V., Rivero Vega, R., Pérez Suárez, R., Centella Artola, A. (2013). Impacto del cambio climático y medidas de adaptación en Cuba. La Habana, Cuba: Instituto de Meteorología, Agencia de Medio Ambiente, Ministerio de Ciencia, Medio Ambiente y Tecnología. Recuperado a partir de http://www.revistaccuba.cu/index.php/acc/article/download/387/318
Rundel, A. (1998). Ecology and biogeography of Pinus: and introduction. En D. M. Richardson, Ecology and Biogeography of Pinus (pp. 3-46). Cambridge.UK, USA: Cambridge University Press. Recuperado a partir de http://mfkp.org/INRMM/article/13504288
Saunders, D. A., Hobbs, R. J., Margules, C. R. (1991). Biological consequences of ecosystem fragmentation: A review. Conservation Biology, 5(1), 18-32. https://doi.org/10.1111/j.1523-1739.1991.tb00384.x
Silva, L., Dias, E., Sardos, J., Azevedo, E., Schaefer, H., Moura, M. (2015). Towards a more holistic research approach to plant conservation: the case of rare plants on oceanic islands. AoB Plants, 7(1). https://doi.org/10.1093/aobpla/plv066
Téllez, V., O., Chabes H, Y. M., Gómez-Tagle, C., A., Gutiérrez G, M. V. (2005). Modelado bioclimático como herramienta para el manejo forestal. Revista de Ciencias Forestales México, 29(95), 61-82.
VanDerWal, J., Shoo, L. P., Johnson, C. N., Williams, S. E. (2009). Abundance and the environmental niche: Environmental suitability estimated from niche models predicts the upper limit of local abundance. The American Naturalist, 174(2), 282-291. https://doi.org/10.1086/600087
Whittaker, R. J., Fernández-Palacios, J. (1998). Island Biogeography: ecology, evolution and conservation. New York, USA: Oxford University Press. Recuperado a partir de https://global.oup.com/academic/product/island-biogeography-9780198566120?cc=cu&lang=en&
Yordani Crespo Martínez. Universidad de Pinar del Río "Hermanos Saíz Montes de Oca". Cuba.